GEEAM
![]()
Les
noyaux gris centraux et le système
sous cortico-spinal ("extrapyramidal")
(Dr. Philippe LECAVORZIN - novembre
2001)
|
|
| COURS |
| ATLAS VIDEO |
1- NEUROANATOMIE FONCTIONNELLE1-1 Description
Le système extrapyramidal est un système moteur polysynaptique, naissant en dehors de l'aire 4 de Brodmann, faisant relais dans les noyaux gris centraux (NGC) ou les noyaux moteurs du tronc cérébral.
Il est constitué des formations suivantes :
-
les aires corticales prérolandiques 6, 8 ; post rolandiques 1, 2, 3, 5 et 22 (Brodmann)
Figure 1.

-
les noyaux gris centaux (NGC) : il s'agit d'un groupe de noyaux sous-corticaux richement interconnectés :
- le noyau caudé
- le noyau lenticulaire, découpé en 2 structures d'origine embryologique différente : une partie externe : le putamen, et une partie interne, le pallidum
- le thalamus moteur par ses noyaux ventral antérieur et ventro-latéral
- des formations sous-thalamiques et mésencéphaliques : le noyau sous thalamique (NST = corps de Luys), le locus niger (= substance noire : SN). La SN comporte 2 parties, la pars compacta (SNpc, composée de neurones dopaminergiques), et la pars réticulata (SNpr, composée de neurones GABaergiques)
- le noyau pédonculopontin (faisant partie de la formation réticulée du tronc cérébral)
Figure 2.
Figure 3.


-
les voies extra-pyramidales prennent naissance dans le tronc cérébral. On distingue 2 groupes de voies descendantes vers la moelle (Kuypers, 1982) :
- le groupe ventromédial chemine dans la moelle ventrale. Il comprend :
. les faisceaux vestibulo-spinaux latéral et médian
. le faisceau tecto-spinal
. les faisceaux réticulo-spinaux
Ces faisceaux se terminent sur la région ventromédiane de la substance grise de la moelle sur des interneurones (IN), des neurones propriospinaux longs et les alpha-MN des muscles axiaux et proximaux des membres. Ils jouent un rôle majeur dans le contrôle de la posture et de l'équilibre.
Figure 4.
- le groupe dorsolatéral chemine dans le quadrant dorsal de la moelle. Il comprend :
. le faisceau réticulo-spinal
. les faisceaux bulbo-spinaux
Ces faisceaux se terminent sur la région dorsolatérale de la substance grise de la moelle sur des interneurones (IN), des neurones propriospinaux courts et les alpha-MN des muscles distaux des membres. Ils sont impliqués dans les mouvements distaux des membres.1-2 Organisation fonctionnelle
Les noyaux gris centraux sont groupés en ensembles fonctionnels :

-
le striatum (striatum dorsal ou néostriatum) comprend 2 structures séparées par la capsule interne : le noyau caudé et le putamen. C'est la partie la plus volumineuse des NGC ; elle possède la même origine embryologique. Le striatum représente la structure d'entrée des NGC. Il est doté de fonctions réceptrices et associatives. A côté de ces noyaux qui constituent la plus grande part du striatum (ou striatum "dorsal"), on décrit un striatum "ventral" (Heimer, 1982) formé du noyau accumbens et de certaines régions du tubercule olfactif, connecté à un pallidum ventral (partie rostrale de la substantia innominata). Striatum ventral et pallidum ventral permettent l'interface entre le système limbique et le système moteur. Enfin, certains considèrent le NST comme une structure d'entrée des NGC, au même titre que le striatum.
-
le pallidum ou globe pâle (globus pallidus)correspond à la partie interne du noyau lenticulaire. Il a un rôle effecteur. C'est la structure de sortie des NGC. On distingue un pallidum externe (Gpe) et un pallidum interne (Gpi)
-
la SNpr est la seconde structure de sortie des NG. Elle se projette sur la formation réticulée et le Noyau Pédonculo-Pontin (fibres nigro-tegmentales), le collicullus supérieur (situé dans le toit du mésencéphale, fibres nigro-tectales), à l'origine de voies descendantes vers la moelle (voies "extrapyramidales" tecto, réticulospinales).
-
le rôle fonctionnel des noyaux gris centraux reste mal connu. Il concerne globalement la programmation motrice et est impliqué dans tous les types de mouvements.
-
il existe dans le pallidum une activité tonique inhibitrice (GABAergique) dirigée vers le thalamus moteur, qui freine l'action de neurones thalamo-corticaux se projetant sur l'aire motrice supplémentaire (AMS), et entrave donc l'expression motrice
-
cette inhibition motrice peut être levée par le striatum, qui projette sur le pallidum des fibres inhibitrices (également GABAergiques)
-
cette désinhibition est obtenue sous l'influence d'afférences corticales excitatrices (glutamatergiques) se projetant sur le striatum. Elle permet de libérer de façon très sélective des circuits moteurs spécifiques (Chevalier et Deniau, 1990), en réponse à des stimuli environnementaux particuliers; elle permet donc de "focaliser" l'activité corticale en sélectionnant au sein d'une activité diffuse, les influx corticaux adaptés au contexte (Houk et Wise, 1995). Ce processus de sélection motrice est soumis à la régulation du locus niger (SN pars compacta - dopaminergique).
-

Figure 5.
-
Le striatum est impliqué dans la programmation de tous type de mouvements, mais en particulier:
-
lorsque la tâche motrice est réalisée sans possibilité de contrôle par des indices sensoriels (mouvement autoguidé). La programmation motrice fait alors appel à des représentations internes (indices internes - internal cues) nécessitant une trace mnésique, en particulier de nature spatiale. Le striatum intervient grâce aux projections nombreuses qu'il reçoit du cortex associatif (les aires motrices se projettent sur le putamen, alors que les aires associatives se projettent sur le noyau caudé: le cortex frontal vers sa tête, le cortex temporal inférieur vers sa queue). Ainsi, le striatum intervient dans l'élaboration, le stockage et le rappel des programmes moteurs.
-
De plus, par ses connections avec les systèmes motivationnels et de récompense (amygdale, système limbique - striatum ventral), le striatum joue un rôle dans l'apprentissage de tâches comportementales dirigées vers un but. Les neurones dopaminergiques de la SNpc interviennent en signalant au striatum la survenue d'un stimulus à traiter de façon prioritaire. L'activité dopaminergique induit dans le striatum et le cortex préfrontal des processus plastiques, qui confèrent à des stimuli initialement neutres, des propriétés motivationnelles.
-
Certains neurones du putamen sont actifs en préparation et au cours de mouvements lents exigeant une fixation posturale, mais aussi un effort mnésique.
-
1-3 Circuits spécifiques
-
l'organisation détaillée des NGC met en jeu des boucles parallèles (Alexander, 1986) ou circuits. Concernant la motricité, il existe un circuit principal et 4 circuits accessoires :
-
le circuit principal est le circuit cortico-striato-pallido-thalamo-cortical. Cette boucle joue, comme on l'a vu, un rôle dans l'initiation et le déroulement normal des programmes moteurs.
-
les circuits accessoires sont essentiellement des boucles de régulation :
. le circuit striato-nigro-strié ou boucle nigro-striée contrôle l'activité du striatum
. le circuit pallido-sous thalamo-pallidal (ou boucle pallido-luysienne) permet un contrôle de l'activité pallidale, le NST recevant des projections corticales directes.
. le circuit striato-pallido-thalamo-strié. Il permet la régulation des informations sensorielles plurimodales provenant du thalamus
. le circuit cortico-striato-nigro-thalamo-cortical -
-
A côté du circuit principal destiné au cortex, qui module l'activité du système cortico-spinal (Jellinger, 1990), le pallidum envoie également des projections vers le locus niger (SNpr), les formations mésencéphaliques comme le noyau pédonculo-pontin (NPP), la substance réticulée et le noyau rouge. Ces formations sont à l'origine des voies tecto, rubro, réticulospinales, qui jouent un rôle dans l'adaptation du tonus postural ou la motricité des membres.

Figure 6.
L'organisation fonctionnelle des noyaux gris centraux.
Circuit principal (1,2,3,5,6). Le N. Caudé et le putamen (striatum) reçoivent des afférences de l'ensemble du cortex, en particulier des aires motrices homolatérales (1). Une voie gabaergique inhibitrice se projette du striatum vers le pallidum (2), et du pallidum vers le thalamus moteur (N. ventral antérieur et ventrolatéral) (3), permettant de libérer l'activité des neurones thalamo-corticaux se projetant sur le cortex moteur et prémoteur (circuit striato-pallido-thalamo-cortical) (5), dont la voie principale de sortie est la voie pyramidale (6).
Il existe plusieurs boucles de régulation de ce circuit principal. La boucle nigro-striée (8) prend naissance dans la SNpc, se projette notamment sur les neurones cholinergiques du striatum par l'intermédiaire de synapses dopaminergiques. Un feed-back inhibiteur (GABA) revient vers la SN, permettant au striatum de moduler ses besoins en dopamine. Le NST échange quant à lui des projections avec le pallidum, à l'origine de la boucle pallido-luyssienne, qui contrôle l'activité du pallidum. Enfin, les noyaux intralaminaires du thalamus reçoivent des afférences régulatrices du pallidum interne, de l'aire motrice corticale (afférences excitatrices), du cervelet et de la réticulée mésencéphalique et envoient des fibres dopaminergiques dans le putamen (9), permettant de réguler l'activité du striatum (boucle pallido-thalamo-striatale).
Le pallidum envoie également des projections descendantes vers le locus niger (SNpr) (4), les noyaux moteurs du tronc cérébral (noyau pédonculo-pontin (NPP), substance réticulée, noyau rouge). Ces formations sont à l'origine des voies extrapyramidales qui contrôlent le tonus postural et participent à la motricité des membres.
- Les circuits moteurs des NGC sont organisés en 2 voies parallèles: 1 voie directe et une voie indirecte, exerçant un effet opposé sur le système moteur (Albin, 1989, Delong, 1990):
- la voie directe prend naissance dans le striatum et se projette, par l'intermédiaire de fibres GABAergiques inhibitrices, sur la partie interne du pallidum (Gpi) et la SNpr, qui à leur tour projettent sur les noyaux moteurs du tronc cérébral et le thalamus. Cette voie exerce des effets facilitateurs sur la motricité. Elle est facilitée par les projections nigrostriées excitatrices mettant en jeu des récepteurs dopaminergiques de type D1.
- La voie indirecte prend naissance dans le striatum et se projette, par l'intermédiaire de fibres GABAergiques inhibitrices, sur le Gpe puis le NST. Celui-ci se projette à son tour, cette fois par des fibres excitatrices glutamatergiques, sur le Gpi. Cette voie exerce des effets inhibiteurs sur le thalamus et les noyaux prémoteurs du tronc cérébral. Elle est inhibée par les projections nigrostriées inhibitrices mettant en jeu des récepteurs dopaminergiques de type D2.
- Le rôle respectif de chacune de ces voies est débattu:
- certains considèrent que le cortex activerait d'abord la voie directe pour initier le mouvement, puis la voie indirecte pour l'interrompre (Alexander, 1990)
- d'autres proposent que le cortex activerait simultanément les 2 voies, la voie directe permettant l'exécution du mouvement, la voie indirecte ayant pour rôle d'empècher l'exécution des mouvements non programmés (Mink et Thach, 1991).
- Les mouvements anormaux résultent d'un déséquilibre entre la voie directe et indirecte. Au cours de la maladie de Parkinson, la carence dopaminergique provoque une hyperactivité du Gpi et une freination accrue du thalamus moteur.
2- SEMIOLOGIE : Prototype = Syndrome Parkinsonien
Syndrome = ensemble de symptômes = entité nosologique
parkinsonien : groupe 3 symptomes essentiels :
* tremblement :
- de repos : il s'agit d'oscillations régulières de fréquence 4-7 Hz, survenant uniquement au repos, s'atténuant lors du mouvement. Il prédomine au membre supérieur, généralement en distalité (main ++). Il est renforcé par les émotions, disparaît pendant le sommeil
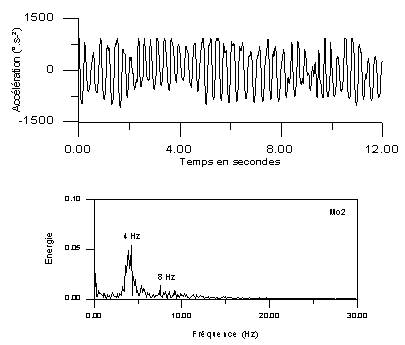
Figure 7.
- d'attitude : il survient lors du maintien d'un membre dans une position donnée
* hypertonie :
- extrapyramidale (hypertonie plastique, continue lors de l'allongement musculaire, non ou peu liée à la vitesse d'étirement, donc différente de l'hypertonie spastique). Elle atteint toute la musculature mais prédomine sur les muscles antigravifiques. Elle peut céder par à-coups, par exeple au poignet, où elle réalise alors le phénomène de la "roue dentée". Cette hypertonie est renforcée par un mouvement du membre controlatéral (signe de Froment). Comme le tremblement, elle disparaît au cours du sommeil)
- exagération des réflexes de posture: c'est l'exagération du réflexe de raccourcissement ("shortening reaction"), bien mise en évidence au niveau du cou de pied (mise en tension du tendon du JA lors de la dorsiflexion du pied)
- l'hypertonie est à l'origine des modifications posturales caractéristiques des patients parkinsoniens (tête et tronc penchés en avant, genoux légèrement fléchis, bras fléchis, collés au corps)* Akinésie :
Elle se manifeste par une rareté du geste, des difficultés d'initiation du mouvement, une amimie (visage peu expressif), la rareté du clignement des paupières, la perte du ballant du bras à la marche, avec une démarche à petits pas. De même, la parole est rare, il existe une micrographie (diminution de la taille de l'écriture).
L'akinésie s'explique par une altération de la motricité autoguidée, sous la dominance du système extrapyramidal, et par l'intervention des systèmes de compensation (motricité facilitée par guides externes) : tout mouvement, pour être réalisé, doit être conçu, programmé, réalisé et supervisé consciemment. Dans l'akinésie, le programme moteur intact, mais il existe un altération de l'exécution des programmes moteurs et de leur enchaînement.
3- FORMES CLINIQUES SELON L'ETIOLOGIE
3-1 Maladie de Parkinson ("paralysie agitante")
Etiopathogénie : dégénérescence du locus niger (pars compacta) avec raréfaction du pigment mélanique. Traitement à base de L-Dopa.
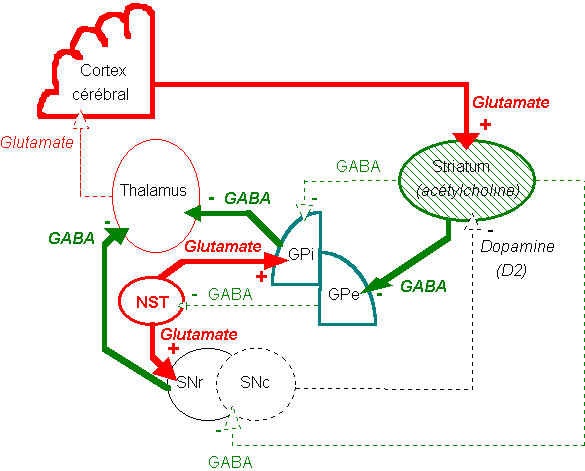
Figure 8.
Tableau clinique :
* Triade symptomatique = akinésie (signe le plus constant) + rigidité + tremblement.
* Signes accessoires : les réflexes ostéo-tendineux (ROT), le réflexe naso-palpébral peuvent être vifs. Il peut exister des signes sensitifs (douleurs profondes), des signes végétatifs, une hypersialorhée, voire des troubles cognitifs.Physiopathologie :
- tremblement : il s'explique par l'hyperactivité d'un réflexe à longue boucle (réflexe "transcortical"), ou une activité rythmique centrale (théorie de "l'oscillateur central")
- akinésie : elle résulte de l'hypoactivité d'un système prémoteur mesial (AMS, gyrus cingulaire antérieur) et du cortex pré-frontal dorso-latéral, intervenant préférentiellement dans les mouvements autoguidés. Par contre, il existe la préservation et le rôle compensateur d'un système moteur latéral (cortex moteur primaire, cortex prémoteur latéral, cortex pariétal et cervelet), intervenant dans les mouvements facilités par un guide externe (Samuel, 1997).
- rigidité : elle pourrait s'expliquer par l'hyperactivité du faisceau réticulospinal dorsal connecté à la SNpr, qui serait responsable des anomalies observées à l'étage spinal (hyperactivité des IN de l'inhibition réciproque Ia, diminution de l'inhibition Ib) selon Delwaide (Delwaide, 19**)
3-2 Autres maladies neurodégénératives (autres signes associés)
- atrohie multisystème (multisystem atrophy, MSA) : regroupe l'atrophie olivo-ponto-cérébelleuse, le syndrome de Shy-Drager
- dégénérescence cortico-basale (DCB)
- maladie de Steele-Richardson (paralysie supranucléaire progressive: PSP)
- voire démences (Alzheimer : autres signes associés)
3-3 Causes vasculaires
Le plus souvent, il s'agit d'un syndrome lacunaire ou pseudo-bulbaire plutôt que d'un syndrôme parkinsonien vrai (hypertonie, rire et pleurer spasmodique, troubles de déglutition, syndrome pyramidal bilatéral). Les lésions sont disséminées dans les noyaux gris centraux (multiples infarctus artériels sur les petites artères vascularisant les NGC). L'évolution se fait par poussées.
3-4 Causes toxiques et médicamenteuses
Intoxication par le monoxyde de carbone (CO) ou le manganèse, les neuroleptiques (signes bilatéraux, symétriques).
3-5 Causes tumorales
Tumeur des NGC ou plus souvent de la ligne médiane ou frontale (action à distance sur la boucle nigro-striée).
3-6 Causes infectieuses
Encéphalite de Von Economo (1917-1925), maladie de Creutzfeld-Jakob (autres signes associés)
4- REFERENCES
- J De Recondo. Principaux syndrômes neurologiques (3ème édition). Roussel Ed, 1980.
- H Ollat. La motricité. Monographies de l'ANPP-volume 4.
- M. Ziegler, J. de Recondo, A. Richer. Maladie de Parkinson et syndromes parkinsoniens. Encycl Med Chir, Neurologie, 17-062-A-10, 1994, 23 p.
- Kuypers HGJM. A new look at the organization of the motor system. In: anatomy of descending pathways to the spinal cord; Eds: HGJM Kuypers, GF Martin. Progr Brain Res 1982, 57: 381-403.
- Heimer L, switzer RD, van Hoesen GW. Ventral striatum and ventral pallidum. Components of the motor system? Tins, 1982, 5: 83-87.
- Chevalier G, Deniau JM. Disinhibition as a basic process in the expression of striatal functions. Trends neurosci 1990; 13: 277-280.
- Alexander GE, Delong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Ann Rev Neurosci 1986, 9: 357-381.
- Albin RL, Young AB, Penney JB. The functional anatomy of basal ganglia disorders. Tins 1989, 12: 366-375.
- Mink JW, Thach WT. Basal ganglia motor control. 3. Pallidal ablation: normal reaction time, muscle cocontraction and slow movements. J neurophysiol 1991; 65: 330-351.
- P Rondot. L'akinésie et les atteintes dissociées du mouvement au cours de la maladie de Parkinson et les lésions des ganglions de la base. Rev neurol, 1990; 146, 10: 591-599.
- M Samuel et al. Evidence for lateral premotor and parietal overactivity in Parkinson's disease during sequential and bimanual movements. A PET study. Brain 1997; 120 : 963-976.
- PJ Delwaide. La rigidité parkinsonienne : aspects cliniques et physiopathologiques. Rev neurol, 1990, 146, 10: 548-554.
- K. Jellinger. New developments in the pathology of Parkinson's disease. Adv Neurol 1990 ; 53 : 1-16.
5- WEB
- P. Le Cavorzin.
Pharmacologie de la maladie de Parkinson.
http://www.med.univ-rennes1.fr/etud/pharmaco/